
汉唐中医倪海厦撰写

临床研究期刊(TheJournalofClinicalInvestigation)Volume117Number5May2007
标题:
以中和抗体抑制肿瘤成长因子β可避免因放射治疗引起的肿瘤加速转移
(InhibitionofTGF-βwithneutralizingantibodiespreventsradiation-inducedaccelerationofmetastaticcancerprogression)
作者
SwatiBiswas,1MartaGuix,2CammieRinehart,2TeresaC。Dugger,2AnnaChytil,1
HaroldL。Moses,1,3,4MichaelL。Freeman,5andCarlosL。Arteaga1,2,4
单位
属于美国田纳西州那什维尔范德堡大学(VanderbiltUniversity)
1。癌症生物学系、2。医学系、3。病理学系、4。Vanderbilt-Ingram癌症中心乳癌研究计划以及5。放射肿瘤学系
摘要
我们探究了因癌症治疗而产生的TGF-β究竟会不会加速肿瘤的发生。我们使用了MMTV/PyVmT此一转移型乳癌基因的克隆模型,指出了游离辐射(译注1)或doxorubicin(译注2)都会增加循环中TGF-β1与肿瘤细胞的量,并会增加肺部的癌症转移。这些效应都因为我们使用了泛TGF-β抗体而受抵销。我们在有TGF-β抗体的环境下,体外培养了这些在循环中会表现多瘤病毒中等T抗原(译注3)的肿瘤细胞。
发现它们不会生长;这表示TGF-β在这些细胞中是一种生存的讯号。当老鼠的肿瘤不含有第二型TGF-β受器时,放射线就不会增加肺部的癌症转移;这显示至少在部分上,肺癌转移是跟TGF-β在肿瘤细胞上的直接作用有关的。这些数据含示,癌症治疗所产生的TGF-β是一种帮助癌症转移的讯号,也提供了一个原理根据,让我们得在癌症治疗时同时使用TGF-β抑制剂。
译注1:此为放射性治疗。
译注2:此为一癌症化学治疗常用药物,俗称“小红莓”。
译注3:此为感染此病毒之初期的产生的三种T抗原中其中一种。
介绍
肿瘤成长因子TGF-β,既是一个肿瘤抑制基因,也是一个肿瘤启动子。当TGF-β配体(ligands)结合到与其对应的丝胺酸/酥胺酸(Ser/Thr)激酶跨胞膜受器后,这些受器接着就会磷酸化、活化Smad族的讯息传递者。被活化之后,Smad2、Smad3会与Smad4结合并迁移至细胞核内,在此处调控与细胞周期停止(cellcyclearrest)、细胞凋亡(apoptosis)相关的基因之转录。
TGF-β之所以有肿瘤抑制基因之用,即系此项调控之功。的确,在内皮细胞与基质中的TGF-β讯息传递,若是丧失或减弱,都会让内皮细胞的细胞恶性转化(transformation)大行其道。另一方面,已有研究显示,若将显性且无效的TGF-β受器引入至移转的癌症细胞中,从上皮细胞型态转变成间叶细胞型态的转分化(epithelial-to-mesenchymaltransdifferentiation)、能动性(motility)、侵略性(invasiveness)、以及存活率(survival)均会受到抑制。
这证明在已经完全转化的细胞中,TGF-β扮演了一个肿瘤启动子的角色(于参考文献4中有回顾)。大部分的癌都保有TGF-β受器,却丧失或部分丧失了Smad依赖性的抗细胞分裂效应(Smad-dependentantimitogeniceffect);同时,在某些个案中,他们还会对TGF-β有所反应,而获得了帮助癌症转移的能力。进而,不论癌症细胞过度生产TGF-β、过度活化TGF-β。
或是两者兼备,都会借由旁泌(paracrine)的方式,改变肿瘤周围的微小环境(microenvironment),帮助肿瘤的发生。这些资料提供了支持根据,让我们得以设法阻断人类癌症中的自泌性(autocrine)或旁泌性(paracrine)之TGF-β讯息传递,以达治疗之目的。
除Smad族的蛋白质外,TGF-β还可以刺激数种转化的讯息传递路径。之前TGF-β已被证实可保护转化细胞(transformedcells)不受细胞凋亡之害。此类细胞反应有一种可能的机制,就是TGF-β诱发了PI3K和PI3K的标的物,也就是丝胺酸/酥胺酸激酶Akt,此讯息传递程序和对于癌症药物的抗药性是相关连的。有些对传统化学疗法有抗药性的肿瘤会过度表现TGF-β。
而TGF-β的抑制剂经证明可逆转此种抗药性。还有,大部分的肿瘤都会过度表现TGF-β配体(ligands)。在肿瘤组织与在血清中的高量TGF-β配体,与肿瘤转移的复发、患者较差的愈后,均有所相关。
在乳癌的基因克隆模型中,TGF-β之讯息传递可加强转移既存的乳腺肿瘤。这是受例如Neu/ErbB2或多腺瘤病毒中央T抗原(polyomavirusmiddleTantigen,PyVmT)等的癌症基因之诱发所致。再者,在MMTV/LTR乳腺启动子的控制下,表现PyVmT癌症基因的基因克隆鼠,经过有条件的TGF-β1诱导,只要两个礼拜之短。
转移肺癌的几率便高了十倍。有些癌症治疗已被证实会全面性地、或留在原处诱发TGF-β。所以我们推测,被癌症治疗所诱发的TGF-β,会提供存活讯息给这些对化疗有抗药性的肿瘤里的细胞,或者一部分此种肿瘤中的细胞,使它们可能在治疗之后,立即加速肿瘤的扩展。在此,我们使用了乳癌转移的MMTV/PyVmT基因克隆模型来展示,游离辐射(ionizingradiation)和小红莓(doxorubicin)提高了循环中的TGF-β1浓度、循环中的肿瘤细胞数、以及肺部的癌症转移。
这些效应都借由TGF-β中和抗体的施加,而得以抵销。在缺乏第二型TGF-β受器(TβRII)的老鼠身上,放射线则没有增加肺部的癌症转移。这些资料暗示,癌症治疗所产生的TGF-β是肿瘤中一个帮助癌症转移的讯号,因此也提供了原理根据,使我们在使用这些疗法时,得以同时搭配使用TGF-β的抑制剂。
结果
胸腔放射线处理与化学治疗提高了循环中的TGF-β量。我们在八周大的未交配母鼠之胸腔与骨盆内,施加了10Gy的放射线。在施加放射线之后隔24小时,进行血液采集。我们在受放射处理之老鼠的血浆里,观察到约为控制组2倍的TGF-β1含量(胸腔,P=0。03;骨盆,P=0。02;图一A);同时,TGF-β2的量没有改变(资料未呈现)。在八周大的MMTV/PyVmT基因克隆鼠中。
或是在种下MMTV/PyVmT肿瘤细胞、稳定地转染了luciferase表现载体的非基因克隆鼠中,我们都得到类似的结果(分别为P=0。015与P=0。007,相较于控制组,图一B)。在经过放射处理后,TGF-β1的浓度有持续七天比控制组高(资料未呈现)。为了将这些结果推广到其他的癌症治疗法,我们测试了DNA嵌入剂(DNA-intercalatingagent)与topoisomeraseII之抑制剂doxorubicin(Adriamycin)的效果如何。
从第八周开始,基因克隆鼠被施行三次腹腔注射doxorubicin(5mg/kgi。p。),每次分别间隔21天。在第十五周收集到的血浆里发现,相较于未处理的老鼠,TGF-β1浓度提高了两倍(P=0。009;图一C),而TGF-β2浓度则是持平不变。为了要测量在放射处理后五周所取得的肺脏组织中,被活化的TGF-β1之浓度,我们使用了一种TGF-β1生物检验来检测,这检测是使用一种会稳定表现plasminogenactivatorinhibitor-1/luciferasereporter(PAI-1/luciferasereporter)的貂肺内皮细胞来运作的。
相较于未受放射处理之老鼠肺组织裂解液,经过放射线处理的老鼠肺组织裂解液,诱导提升了有2倍多的TGF-β1浓度(P=0。0008;图一D)。
胸腔的放射线处理及化学治疗,会增加循环中的肿瘤细胞量与增加肺部癌症转移。TGF-β增强了会表现PyVmT肿瘤基因之肿瘤细胞的存活率,而且之前已被证实,TGF-β有条件地被诱发后,会在MMTV/PyVmT基因克隆鼠中加速乳腺肿瘤的转移。因此,我们推测,因治疗而提升的循环中TGF-β1浓度,可能和这个乳癌转移模型中的循环中肿瘤细胞增加、肺部癌症转移增加。
有所关连。带有肿瘤的MMTV/PyVmT老鼠在第八周大时,被施行胸腔放射处理(thoracicirradiation)(10Gy)直至第十三周为止。实验终结时进行心脏穿刺以取得血液,而血液中的细胞部分,则拿去测试其在活体外生成细胞集落之能力。从未经放射处理之老鼠所取得之血液,产生了平均1。6±1。5个细胞集落;而从经过放射处理之老鼠所取得之血液。
于10-12天后所测量的结果是,产生了平均28。3±7。6个细胞集落(P=0。02:图二A)。PyVmT抗体的免疫染色证实,这些生长的细胞集落有表现癌症基因(图二A)。循环中的活肿瘤细胞数有所增加;而和此相符合的是,我们在经放射处理的老鼠中,观察到较未经处理之控制组17倍的肺部表面癌症转移(P=0。009;图二B)。在被用doxorubicin处理相较于未处理之老鼠中。
也在肺部癌症转移观察到类似的上升情形(P=0。032;图二C)。在施加doxorubicin24小时之后,相较于未经处理之控制组老鼠所取得之血液,也观察到在细胞群落数目的上升(17。2±3。7与7。2±2colonies;n=5;P=0。007)。从老鼠的体重及一系列的肿瘤直径数据来看,受处理与未处理的老鼠在肿瘤负荷量上并无差异。
这表示TGF-β1浓度之差别,不能简单地以肿瘤负荷量会随时间增加来解释。
接下来,我们检验了放射线所诱发之肺部癌症转移,是否仅限于带有肿瘤的MMTV/PyVmT基因克隆鼠身上。为了测试这个可能性,我们从会表现PyVmT之基因克隆的乳腺癌中,取得表现luciferase的肿瘤细胞,然后将其注射至非基因克隆的同源FVB老鼠身上。这些老鼠中的肿瘤,生长在左侧鼠蹊(inguinal)(四号)的乳腺脂肪垫,我们以生物发光法,观察并估计了其转移扩散至肺脏的情形(图三A)。
当肿瘤长到了200mm3或更大时,老鼠在胸腔施加了10Gy的放射线并在其余部分施加屏障,两周后将老鼠牺牲。相较于控制组,接受放射处理的老鼠增加了三倍的表面肺部癌症转移(P=0。005;图三,B和C)。再者,在放射处理后24小时与两周之后,分别有相较于控制组多8倍与5倍的循环中肿瘤细胞量(表一)。在实验组与控制组间,原发肿瘤的生长率并无不同(资料未呈现)。
将培养中的PyVmT细胞暴露在1。25-7。50Gy下,会增加其产生TGF-β的量(图四A)。在以上所述之研究过程中,有既成之肿瘤的老鼠才会施加放射线处理;循环中TGF-β浓度的增加,有可能是因为放射线在癌症细胞上的作用,因此我们想排除掉这个可能。我们对未交配的FVB母鼠施加了10Gy的胸腔放射。在放射处理后一个小时,我们从尾部静脉注入了表现PyVmT且受luciferase稳定转染(stablytransfected)的肿瘤细胞。
肺脏中的肿瘤细胞负荷量(tumorcellburden),是在肿瘤细胞的注入之后,使用活体内(invivo)与活体外(exvivo)的生物发光来监测的。经放射线处理的老鼠肺脏,表现了较控制组为高的生物发光讯号(图四B),此讯号与人工计数之表面肺癌转移数量是相关连的,亦与肺脏重量相关(图四,C与D)。
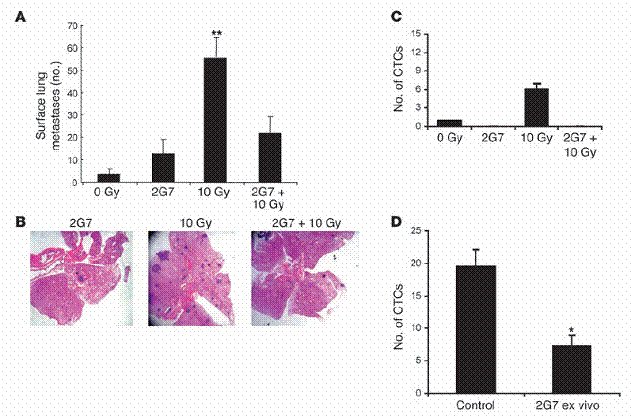
TGF-β中和抗体阻断了受放射线诱导增加之肺部癌症转移。我们使用了2G7中和泛TGF-βIgG2抗体(2G7neutralizingpan-TGF-βIgG2)来验证,放射线所诱导增加的循环中TGF-β浓度,是否在肺癌转移中扮演一个成因的角色。这个单株抗体阻断了所有三种哺乳类所具有的TGF-β异构物,并且在活体中有活性。八周大的MMTV/PyVmT基因克隆鼠。
在胸腔放射处理的2小时之前,以15mg/kg之2G7或PBS注射至腹腔中。这些处理持续每周两次,到第十三周为止,此时将老鼠牺牲并测量是否肺部有癌症转移。然而有趣的是,施打2G7到未经放射线处理的老鼠中,少许增加了肺癌转移,但在统计上并不显著(图五A和B)。在两种实验组别之间,并未存在原发肿瘤细胞负荷量的差异(资料未呈现)。
我们收集了血液并体外培养其细胞悬浮液,以估计在循环中的肿瘤细胞量。从未经放射处理与经过放射处理的老鼠身上,血液中取得而培养的细胞群落数,均因施加2G7而显著地减少(图五C)。最后,我们检验了在体外添加2G7以抑制TGF-β,是否会阻止老鼠血液中取得之肿瘤细胞群落生长。将带有肿瘤、经过放射处理的老鼠所取得之血液,接种于含有2G7与不含有2G7的环境下。
而在十天之后评估其集落生长情形。将细胞培养在存有2G7的环境下,发现会降低70%的细胞群落数(图五D),这表示自泌性的(autocrine)TGF-β对于表现PyVmT的循环中癌症细胞,是一种存活因子。
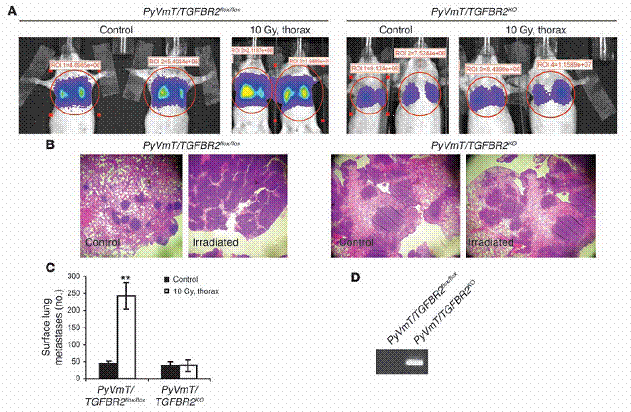
放射线诱导增加肺部癌症转移,需要肿瘤细胞内的TGF-β受器。以上描述的结果显示,因放射线诱导之循环中TGF-β浓度提升,是对于循环中肿瘤细胞的存活信号,让细胞可以移居至肺脏、在肺脏里生长。因此我们推论,借由移除细胞对循环中TGF-β的反应,以使得TβRII不存在于MMTV/PyVmT肿瘤中,就可以抵销放射处理在癌症转移上的效应。为检验此推论之可能。
我们使用了表现PyVmT的肿瘤细胞,并且用Cre-Lox技术将TβRII有条件地于细胞中移除。含有或未含TGFBR2的PyVmT表现型肿瘤细胞株(在此之后将称为PyVmT/TGFBR2flox/floxorPyVmT/TGFBR2KO)是分别从PyVmT/TGFBR2flox/flox及PyVmT/TGFBR2KO老鼠身上的乳腺癌所产生。
PyVmT/TGFBR2flox/flox及PyVmT/TGFBR2KO细胞株均以luciferase之载体稳定转染,且经由尾部静脉注射入八周大、未交配的FVB母鼠,这些母鼠有些没有经过放射线处理,有些则是在注射肿瘤细胞之前1小时,施加了10Gy的胸腔放射。两周后估计肺部癌症转移情形。在被注射PyVmT/TGFBR2flox/flox细胞的老鼠里,经放射处理之老鼠相较于控制组老鼠。
发现生物发光讯号有所增加、肺部癌症转移增加6倍;但如果老鼠之前是被注射不含有TβRII的细胞,在施加与未施加放射处理的两组中,便没有差别(图六,A-C)。PyVmT/TGFBR2KO的肿瘤结节比PyVmT/TGFBR2flox/flox的肿瘤结节还大,此结果与之前的一项报告是相符合的,也就是说,PyVmT表现型乳癌细胞之TβRII丧失,会增进肺部癌症转移的生长。经PCR扩增肿瘤组织中取得之DNA证实了在PyVmT/TGFBR2KO细胞中。
是存在基因重组的(图六D)。
讨论
我们在基因克隆老鼠的乳癌转移模型中,研究了癌症治疗所诱发的TGF-β是否会加速肿瘤的发生。在MMTV/PyVmT基因克隆鼠中,在肺部施加游离辐射或是以doxorubicin处理,都增加了循环中的TGF-β1量、循环中肿瘤细胞数以及肺部癌症转移。循环中TGF-β1浓度增加、之后的癌症转移增加,并不需要老鼠在施加处理时就拥有肿瘤,就如同我们也在放射处理后才施打肿瘤细胞的老鼠中所观察到的。
TGF-β1会增加,并不仅限于受到胸腔放射处理的诱导,因为我们在骨盆的放射处理、以及全面性施加doxorubicin下,也都观察到了TGF-β1的受诱导增加。在放射处理后,循环中肿瘤细胞的增加以及肿瘤转移的增加,都因到2G7的施加而抵销。重要的是,循环中PyVmT表现型肿瘤细胞无法在有2G7的环境下进行体外培养,这也就表示了自泌性(autocrine)TGF-β对这些细胞是一种存活的讯号。
这些结果与之前两项研究是相吻合的;在这些研究中,以可溶性TβRII:Fc融合蛋白、或用反义TGF-β1(antisenseTGF-β1)稳定性转染加以阻断TGF-β,都抑制了肿瘤细胞的能动性、存活率、渗入(intravasation)以及肺部癌症转移。
老鼠身上的肿瘤不含TβRII,放射线便无法在这些老鼠身上加强肺部癌症转移。为了展示此点,我们从PyVmT/TGFBR2flox/flox与PyVmT/TGFBR2KO之老鼠身上既成之乳腺肿瘤,取得细胞株来使用。Forrester等人曾经提出报告,以MMTV/Cre表现有条件地使TGFBR2缺失时,这些knockoutPyVmT表现型的肿瘤发生潜伏期就变短了许多,并且表现出明显增加的肺部癌症转移。
这是相较于PyVmT/TGFBR2flox/flox型的肿瘤。因为我们使用了静脉注射法,并且,我们所注入细胞株是从既成之含有TβRII与未含TβRII的肿瘤而来,所以我们的结果无法解释Forrester等人所报导之癌症潜伏与转移的表现。尽管我们在经放射处理的老鼠中无法排除TGF-β之增加,是否会影响宿主的微环境(microenvironment)与/或免疫系统。
并进而帮助了肿瘤转移之进行,我们不含有TβRII之肿瘤的结果却强烈地显示,癌症转移之增加至少在部分上,是肇因于TGF-β在癌症细胞的直接作用。
在肿瘤细胞中,这些潜在受循环中TGF-β浓度调控的讯号反应,目前仍属未明。然而,TGF-β的助癌转移效应并不仅限于此基因克隆模型。举例说明,在MMTV/LTR启动子控制下表现有活性的TGF-β1与Neu的双基因型(bigenic)的老鼠,也表现出比MMTV/Neu更多的循环中肿瘤细胞与肺部癌症转移。会表现Neu和具活性的TGF-β1之乳腺肿瘤细胞,能侵入Matrigel并且能在transwells上移动。
但却受可溶性的TβRII:Fc给阻断,表示单单Neu不足以诱导出一种侵略性的表现型。再者,在会表现具活性Neu的基因克隆肿瘤里,若共同表现具活性的突变种Alk5和TβRI,会增加肺部血管外的癌症移转(extravascularlungmetastasis),这与TGF-β之效应相符:也就是肿瘤周围蛋白酶、癌症细胞的附着和侵略。PyVmT是一个强而有力的癌症基因。
我们也已经知道它会活化ErK和PI3K;但2G7竟然能对如此的肿瘤细胞具备有效的抗肿瘤移转效果,有点不符合直觉。但是,这表示TGF-β可以放大癌症基因的讯号,而只要这讯号大于某一阈值,可以完全地使肿瘤移转被观察到;并且,相反地,在肿瘤基因已恶性转化的细胞(oncogene-transformedcells)内阻断TGF-β讯号时,会把这些讯号降低至比阈值更低的程度,使其无法到达癌症基因诱导癌症进行。
确实如此,在鳞状细胞癌(squamouscancers)中被逼着表现的显性-具活性Smad2,已经被证实会和有活性的Ras共同作用,使不具侵略性的肿瘤转变成会转移的肿瘤;但Smad突变型不与TβRI结合,并会抑制乳癌细胞的转移。最后,在Ras经恶性转化的细胞中,显性-不具活性且被截短的TβRI表现之后,则是会抑制肿瘤的产生能力(tumorigenicity)以及癌症移转。
癌症治疗后的肿瘤再增殖(repopulation)与发展,是一众所皆知的现象。它已被证实会在放射治疗、化学治疗以及手术摘除后发生。(在参考文献37中有回顾)。TGF-β的浓度会对放射线有所反应而增加,也被发现与放射治疗后之肺脏受伤有关,这是由胶原蛋白沉积、肺泡壁的增厚以及内皮细胞受伤所致。放射线引起的肺脏组织伤害,会因为施以抗TGF-β抗体而显著地减少。在手术方面,已经有人提出。
因为组织操作以及伤口复原而产生的生长激素过量释放,包括TGF-β的过量释放,会紧接着手术之后促进癌症移转。确实如此,手术之后,循环中随即发现了肿瘤细胞的存在,也发现肿瘤细胞数量的增加;而且在治疗性切除直肠癌两周之后,若循环中的TGF-β浓度居高不下,就表示癌症会很快地转移至肝脏。相反地,在手术刚切除直肠肿瘤后,血清中的TGF-β浓度却是显著地降低的。
最后,以化学-放射治疗处理后期的头颈部癌症,以及以化学治疗处理非小型细胞肺癌(non-small-celllungcancers)的情形下,循环中的TGF-β浓度已被证实和对治疗的反应相关。
在本研究中,我们展示了放射治疗与化学治疗会提升循环中的TGF-β浓度、循环中的癌症细胞数、以及癌症的移转。这些效应会因为系统性地施加2G7而被阻断,因此支持TGF-β为癌症转移进展之肇因的说法。这些数据有数个临床上的含意。第一,因癌症治疗而增加的循环中TGF-β浓度,应该要预前地探讨与监控,因为它可能是一种标记,代表肿瘤注定要在治疗之后快速地增长。第二,在循环中TGF-β浓度已被观察到增加的病人。
其身上带有的肿瘤,可能会因我们施加TGF-β抑制剂而加强基本疗程的效果,也可能可以抵销与疗程相关的毒性效应,例如放射线引起的组织伤害以及纤维化。我们预期,以我们现存的方法以及尚在临床研发中的治疗用TGF-β抑制剂,这些假说在不久的将来可以被证实。
方法
药剂与细胞株。2G7融合瘤是B。Fendly(Genentech)所赠。它是在VanderbiltUniversityMolecularRecognitionCorefacility被制出以及用亲和纯化法所得到的。重组的人类TGF-β1以及为TGF-β1用的QuantikineElisakit是从R&DSystems取得。
表现PyVmT的细胞株是从MMTV/PyVmT基因克隆鼠的乳腺肿瘤中取得。PyVmT/TGFBR2flox/flox及PyVmT/TGFBR2KO细胞株是分别从PyVmT/TGFBR2flox/flox及PyVmT/TGFBR2KO老鼠的乳腺癌中取得。我们在这些老鼠中使用Cre/Lox技术,有条件地从乳腺中删除了TGF-βII。细胞被培养在一个37°C、湿化5%的温箱中。
并且培养于添加了10%FBS(HyClone)的DMEM(Cellgro;MediatechInc。)中。
TGF-β1的生物检测。貂肺脏内皮报告细胞(minklungepithelialreportercell)在PAI-1启动子控制下,稳定地表现萤火虫luciferase。这些细胞接种于12-well培养皿上(2x105/well)并且留置过夜供其贴附。已知浓度的可溶性TGF-β1或肺脏组织的细胞裂解液(250μg/well)加入了三次,而这些细胞被留在5%二氧化碳、37°C的环境下培养了另外24小时。
在清洗之后,这些细胞被收取至200μg/well的裂解缓冲液中(DualLuciferaseKit;Promega),luciferase的活性则是在依据厂商提供的实验步骤下进行了测量。
DNA萃取与聚合酶连锁反应(PCR)。以石蜡固定的肿瘤切片被取出和重新添加水分。趁着还潮湿的时候,肿瘤部分以经消毒之刀片刮取至经消毒的eppendorf管中。DNA萃取是使用Instagene(Bio-Rad)并依厂商提供的实验步骤所完成。DNA样本储存在-20°C,除非要另外使用。Cre诱导的PyVmT/TGFBR2KO肿瘤重组,是用PCR引子以及前述之反应条件下所验证的。
眼窝后(retroorbital)血液收集以及TGF-β1之定量。老鼠以1%-2%isofluorane麻醉。血液(约250μl/老鼠)采集于眼睛里的结膜静脉,作法是使用肝素化之Natelsontube(FisherScientific)然后再移转到肝素化的(heparinized)玻璃管中。血浆的制备是使用Ficoll-PaquePlus(AmershamBiosciences)并依照厂商之使用说明进行。
且增加了一个步骤,即在4°C下以10000g离心十分钟以除去血小板。为测量PyVmT表现型细胞之培养基中的TGF-β1,1x106的细胞被涂布在含在完全培养基的10mm盘上,并且供其贴附。该培养基在隔天被更换成无血清之培养基。隔夜培养后,细胞被施以1。25-7。5Gy。过72小时后,培养基经收集并以快速真空法浓缩(3mlto500μl)。
老鼠的血浆和受细胞条件培养基(cell-conditionedmedium),均在下一步的TGF-β1的QuantikineElisakit(R&DSystems)中接受测试,依照的是厂商提供的实验步骤。标准曲线以31。5–2,000pg/ml的人类重组TGF-β1绘出后,被用来计算TGF-β在老鼠血浆与细胞培养基中的等量。
每项标本都在另外的重制实验中重复检验了三次。
TGF-β2之定量。依照厂商提供之实验步骤,在酸活化之后,血浆标本以TGF-β2QuantikineElisakit(R&DSystems)测试。标准曲线以31。5–2,000pg/ml的人类重组TGF-β2绘出后,被用来计算TGF-β2在老鼠血浆的等量。每项标本都在另外的重制实验中重复检验了三次。
老鼠的放射处理。MMTV/PyVmT基因克隆老鼠或正常的FVB母老鼠(Harlan),在麻醉后把背部固定在镜台上。我们使用了单一的胸前辐射场,并使用了铅块掩护老鼠身体的其他部分。在某些实验中则是使用了一个单一腹部辐射场。老鼠被施加300kVp的X光,其剂量为2。05Gy/min。所有的老鼠都依照此法规InstitutionalAnimalCareandUseCommitteeofVanderbiltUniversityMedicalCenter。
被留在一个专门的无病原空间中。在动物暴露在放射线或其他处理后,喂食正常食物,并且详细观察是否有任何情绪不稳的迹象。
小红莓(doxorubicin)的处理。53天大的MMTV/PyVmT老鼠,经腹腔注射三次溶解于食盐水液中的doxorubicin(Adriamycin;5mg/kg;Sigma-Aldrich),每次间隔21天,直到第95天止。经无效空白试剂或小红莓处理之老鼠,其血液样本和肺脏组织,都于第107天解剖取得。为了要检查小红莓处理之后的循环中肿瘤细胞,经无效空白试剂或小红莓处理之老鼠,其血液在取得24小时后。
又加入了单一剂量(5mg/kg)。
侦测循环中肿瘤细胞。循环中肿瘤细胞之培养方法如前述,仅有少部分之修改。扼要地说,血液取得于心脏穿刺。细胞悬浮液(包括buffycoat以及红血球)从血浆中分离出后,接种在6-well盘上,这些盘上布有了生长激素减量之Matrigel(BDBiosciences),并添加有DMEM、10%FBS,之后便于37°C、5%二氧化碳之温箱中培养。隔天,在每个well使用了红血球移除缓冲液(4。
15gNH4Cl,0。5gNaHCO3,0。0186gdisodiumEDTAin200mlwater)以及PBS,数次缓和的冲洗。DMEM与10%FBS(3ml/well)添加后,新鲜的培养基每三天将会补充一次。过了10-12天后,以人工计数大小为50μm以上之细胞集落。PyVmT表现是用免疫细胞化学方法来侦测的。
扼要地说,吸出培养基、以PBS清洗wells,然后细胞集落以10%中性缓冲的福尔马林(formalin),在室温下固定30分钟,再用PBS清洗2次,再于室温下与PBS中之3%BSA共存1小时。培养盘接着培养隔夜,于4°C置放于含有生物素标记的抗PyVmT的老鼠单株抗体(diluted1:500,BIOT-115L;Covance)。然后,加入与streptavidin结合之萤光二级抗体(OregonGreen488。
Invitrogen)室温下1小时,之后使用Hoechst细胞核染色法(1μg/ml十使用分钟)。免疫萤光是以LeicaDMIRB倒置显微镜所观察的。
生产反转录病毒载体以及细胞转导。MMTV/PyVmT细胞,是以luciferase之载体稳定转染的。一个luciferasecassette从pGL3-Basic(Promega)裁切下来,并嵌入pMSCV-puro(Clontech)之多重复制位上。amphotropicpackagingPhoenixcells转染后,产生感染性病毒颗粒。
如前述。培养生长中(subconfluent)之癌症基因表现细胞,在4μg/mlPolybrene(Sigma-Aldrich)中,以病毒上清液转导6小时,而含有病毒颗粒的培养基在6小时之后更换;在48小时后,再添加含有10%已经加热变性之FCS的DMEM,并于7-10天后将抗药的细胞集落合并。
肿瘤产生能力以及肿瘤转移之研究。在1%–2%isofluorane之麻醉下,MMTV/PyVmT/Luc细胞(2×106cellsin200μlPBS)被注射于FVB母鼠的左侧四号乳腺脂肪垫。扼要地说,在注入肿瘤细胞之前,脂肪垫便已经以手术使其暴露在外,而伤口以干净夹子密合,并于10天之后取走。每周以生物发光法检查肿瘤两次。
并系列地以测径器测量之。这些肿瘤的以立方毫米(mm3)计算之体积,是以此数学式计算:v=(w2×l)/2,v代表体积,w代表宽度,l代表长度。当肿瘤达到了200mm3或更大的体积(约两周时间),老鼠就被施以放射线处理,或是不作任何处理。老鼠继续每周两次接受生物发光法的监控观察,直到实验终结。两周之后,将老鼠牺牲并取得基本的肿瘤与肺脏。以解剖显微镜计算肺脏表面之癌症转移数。
之后这些肺脏以10%中性缓冲之福尔马林(formalin)固定之。固定好的肿瘤切片,以H&E染色之,并以显微镜检查是否有肺癌之转移。血液是由心脏穿刺取得以如前述,测量TGF-β之浓度以及循环中之肿瘤细胞数。
在其他的情形里,我们使用了MMTV/PyVmT、PyVmT/TGFBR2flox/flox或PyVmT/TGFBR2KO细胞来表现luciferase。2。5×106之单一细胞悬浮液制备后,重新溶于无菌之PBS液中。0。5×106(in200μl)的细胞被注射入FVB母鼠之侧面尾部静脉中。静脉注射24小时后,肿瘤细胞于老鼠肺部之位置,以生物发光法确定之,并且自此之后每周追踪两次。
为了要在活体内阻断TGF-β,在把10Gy注入老鼠胸腔的前24小时,将15mg/kg的2G7注入老鼠腹腔内,每周两次,持续进行,直到老鼠死亡。
生物发光法显影技术。要将生物发光之讯号与肿瘤负载量建立定量的对应关系,先以溶解于去离子水的luciferinsubstrate(d-luciferinpotassiumsalt;Promega)进行腹腔注射(0。15mg/gbodyweight),在注射之后12分钟内取得影像。根据我们之前的时间记录之实验,在此时间记录讯号高峰处之强度(资料未呈现)。
在取得影像时,将老鼠以isofluorane麻醉。生物发光显影是以VanderbiltUniversitySmallAnimalImagingCenter的IVIS-200显影系统(Xenogen)完成的。我们采用了3-5分钟的整合计算时间,并使用on-chipbinningof8,以增加讯号-噪声比。生物发光显影之定量分析是以LivingImagesoftware(Xenogen)完成。
我们在此定义了感兴趣区域(regionsofinterest,ROI)并测量了integratedphotoncounts。
统计。所有的实验数据均以双尾Student’sttest分析。P值小于0。05被视为显著。
表一
稳定表现luciferase的MMTV/PyVmT细胞,被注射入FVB老鼠的右侧鼠蹊乳腺脂肪垫中。拥有200mm3以上肿瘤的老鼠,不加以放射处理,或是在胸腔施加10Gy的放射处理。在24小时及2周后,以心脏穿刺取得血液,并测量细胞悬浮液在活体外形成细胞集落的能力,如“方法”文中所示。数据为两个分别的实验中,每组五只老鼠的平均值±标准差。RT是指放射治疗(radiationtherapy)。
在同一时间点下,AP=0。016versus0Gy。
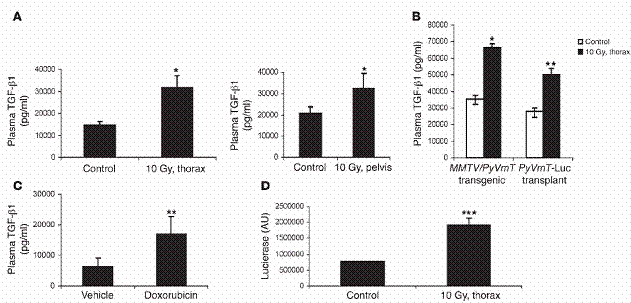
图一
放射线与化学治疗增加循环中TGF-β1浓度。(A)10Gy的放射线被施加于FVB老鼠之胸腔(thorax)(左)或骨盆腔(pelvis)(右)。24小时之后取得血液,血浆中之TGF-β1浓度以在“方法”中所述测量。(B)八周大、带有肿瘤之MMTV/PyVmT老鼠,或非基因克隆之FVB老鼠,在四号乳腺脂肪垫带有200mm3以上大小之肿瘤。
不作任何处理或施加10Gy于胸腔。血浆中TGF-β1浓度,24小时后测量之。(C)在第八周起,每隔21天,以空白试剂或腹腔注射小红莓(5mg/kgi。p。),处理基因克隆老鼠三次。血浆中TGF-β1浓度在第十五周测量之。A–C之数据代表三个不同的独立实验,每组使用三个实验对象。(D)FVB老鼠胸腔被施加10Gy。五周后,取得经放射线处理、控制组老鼠之肺脏。
及其细胞裂解液(250μg/ml),加到含有貂肺上皮细胞的triplicatewells,这些细胞会稳定地表现plasminogenactivatorinhibitor–1/luciferasereporter(PAI-1/luciferasereporter)。二十四小时后,测量luciferase之表现,方法如“方法”部分中所述。图示:*表示P<0。05,**表示P<0。
01,***表示P<0。001相对于控制组。
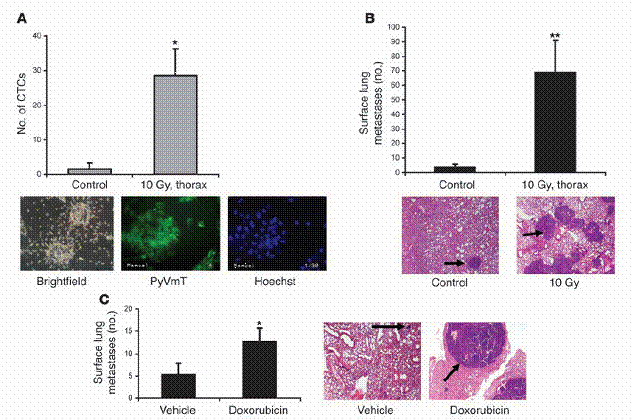
图二
放射线与化学治疗增加循环中肿瘤细胞量以及肺部之肿瘤转移。(A)MMTV/PyVmT母鼠在八周大时,接受胸腔放射线处理。在第十三周时,当实验终结,以心脏穿刺取得血液,并量取其细胞悬浮液在体外产生细胞集落之能力,如“方法”中所述。CTCs代表循环中肿瘤细胞(circulatingtumorcells)。在下面为一些具有代表性的影像,是由血液中取得、循环中单一个肿瘤细胞形成的细胞集落。基因克隆成功的细胞集落。
用PyVmT抗体和萤光之次级抗体处理,并以萤光显微镜观察并估计之。(B)与A同样之老鼠,在第十三周大时计算肺部表面癌症之转移数目。A和B中之数据是代表三个独立的实验,每组中有四只老鼠。从控制组以及经放射线处理之基因克隆老鼠上,取得具代表性之H&E染色的肺部组织切片。这些切片是胸腔放射处理后五周所取得,如下方所示。黑色箭头所指为肺部癌症转移。(C)八周大、带有肿瘤的MMTV/PyVmT基因克隆老鼠。
每隔21天以空白试剂或腹腔注射小红莓(5mg/kgi。p。)处理共三次。此实验于第十五周时终结,并计算肺部表面之癌症转移数量。具代表性之H&E染色肺部切片,其含有癌症转移部分的中心如右方图示。原始放大倍率为100倍。*表示P<0。05,**表示P<0。01,相较于控制组。
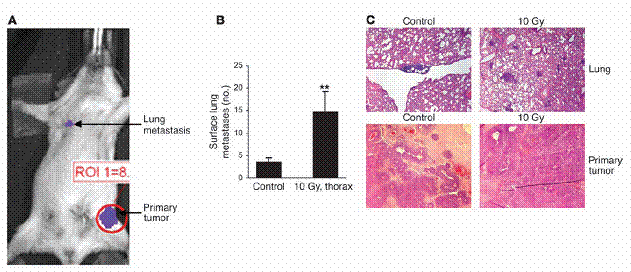
图三
胸腔放射线会增加殖入肿瘤至肺部的癌症转移。(A)稳定表现luciferase的MMTV/PyVmT细胞,被注入同源FVB老鼠之乳腺脂肪垫中。之后,两周一次,以生物发光显影法监测肿瘤生长以及肺部癌症转移。图示为一具代表性之老鼠,图为细胞注入两周之后所摄(B与C)。带有至少200mm3PyVmT/Luc肿瘤的老鼠,以胸腔放射线(10Gy)处理之或不施加任何处理。(B)肺部癌症转移在两周之后定量。
数据为每两次实验中之一次,每组五只老鼠之平均值±标准差。**表示P<0。01相较于控制组(C)肺部与原发肿瘤之H&E切片。原始放大倍率为100倍。
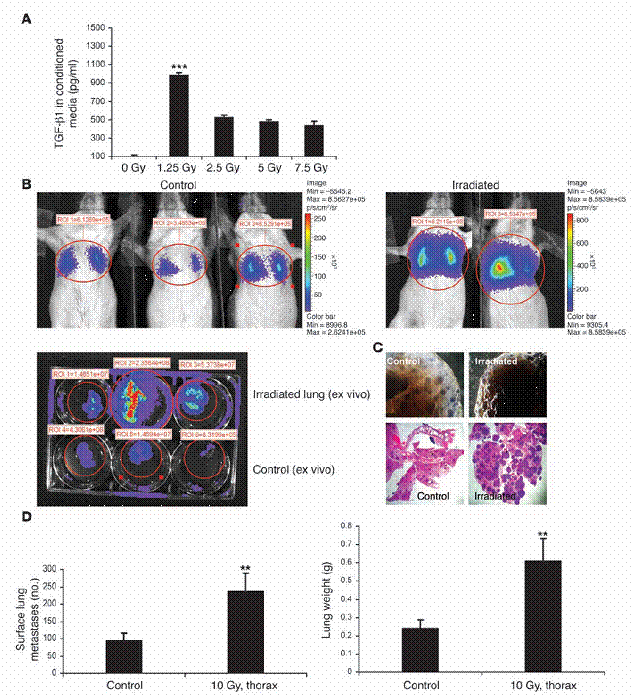
图四
放射线的先行处理,会使移转的肺癌细胞更能够拓展在原本无肿瘤的老鼠身上。(A)MMTV/PyVmT细胞,在加有不含血清之培养基之100-mm培养皿里,以1。25–7。5Gy处理。受细胞所影响之培养基在72小时后进行收集,并以如“方法”中所述之ELISA得知TGF-β1之浓度。(B)稳定表现luciferase之MMTV/PyVmT细胞。
由尾部静脉注入未交配之FVB母鼠。在接受细胞注射1小时之前,老鼠在图中所标部分之胸部,以10Gy处理之。在注射之后2周,肺部之癌症细胞以老鼠的生物发光法进行视觉观察(顶图)。某些情形中,肺脏在添加d-luciferin之后被手术移除,并于体外进行显影(底图)。控制组如左图所示,受放射线处理之老鼠如右图所示。(C)具代表性的整体肺部载片(顶图)以及控制组与放射线处理组之H&E肺部切片(底图。
原放大倍率为100)。(D)在控制组与放射线处理老鼠中,肺部表面癌症转移之定量(左图)与肺部重量(右图)。数据为两个独立实验中,每组的五只老鼠的平均值±标准差。图示:**表示P<0。01,***表示P<0。001,相较于控制组。
图五
TGF-β中和抗体2G7,阻断了放射线诱导增加的肺部癌症转移。(A和B)八周大、带有肿瘤的MMTV/PyVmT老鼠于胸腔接受10Gy处理。在图中标出之部分,老鼠被每周两次施以15mg/kg的2G7,直到第十三周。此时进行肺部表面之癌症转移计数。(A)数据为每组中五只老鼠的平均值±标准差。(B)代表性的H&E染色肺部切片。此实验重复一次取得了类似的结果。
(C)当实验终结,以心脏穿刺取得血液,并量取其细胞悬浮液在体外产生细胞集落之能力,如“方法”部分中所述。(D)在第十三周,经过胸腔放射处理、带有肿瘤的基因克隆鼠,收集血液。细胞悬浮液如C图中情形,在20μg/ml2G7或PBS环境下,涂布于培养盘上。10-12天后,人工计数等于或大于50μm的细胞集落。数据为每组五只老鼠的平均值±标准差。
*表示P<0。05,**表示P<0。01相较于控制组。
图六
肿瘤细胞中不存在TGFβRII,能抵销受放射线诱发的肺部癌症转移之增加。(A–C)未交配的、八周大的FVB母鼠,从尾部静脉注入稳定表现luciferase的PyVmT/TGFBR2flox/flox与PyVmT/TGFBR2KO细胞。这些老鼠在注射肿瘤细胞之前,一组接受、一组未接受胸腔的10Gy处理。两周后,以生物发光(A)、组织染色(B)、以及人工计数肺部表面癌症转移(C)的方法。
来测定肺部表面癌症转移,如“方法”中所述。资料为每组中四只老鼠的平均值±标准差。(D)两组细胞株的基因体DNA之PCR结果,显示重组片段的仅存在于PyVmT/TGFBR2KO细胞中。**表示P<0。01相较于控制组。
译者:JeffreyChuatHsinchu,Taiwan。2009/4/4
不免有所遗漏、错误,敬请参考与指正,谢谢。
文章出处:汉唐中医官网
原文链接:https://www.hantang.com/chinese/ch_Articles/cancer98.htm
免责声明:本站引用或原创的文章,著作权皆属原创作者所有。内容仅限于个人学习和研究使用;禁止将内容用于商业或非法用途,否则后果由用户承担。访问和转载本站内容即表示您已同意上述条款。如有侵犯您的相关权益,请给我们发送邮件:nhx-icu@outlook.com,我们收到后会第一时间删除或者下架内容。

文章评论